Ligenende (strukturelle og funksjonelle) trekk -forts.
(Fra boka 'The Design of Life' kap 5.5-8; av W. Dembski og J. Wells)
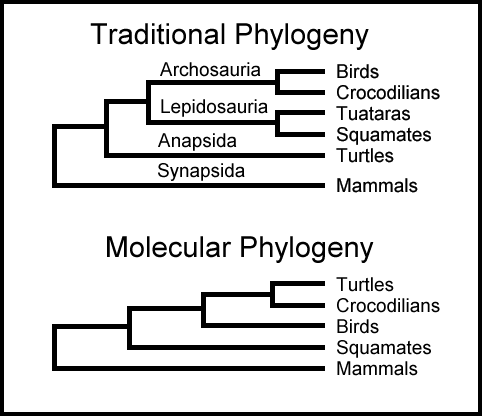 5. Molekylære fylogenier
5. Molekylære fylogenier
En fylogeni er en formodet evolusjonær historie til en gruppe organismer. Inntil 1980 ble fylogenier sluttet ut fra anatomiske og fylogenetiske trekk (benstrukturer, varmblodighet etc.) Med ankomsten til moderne molekylær biologi, er mange fylogenier nå basert på DNA og protein-sammenligninger. Gjennom reproduksjon blir sekvenser av underenheter kopiert fra ett DNA-molekyl til ett annet, men molekylære uhell eller mutasjoner gjør noen ganger kopier litt annerledes enn foreldre-molekylets. Dermed kan organismer ha DNA molekyler (og dermed proteiner) som er litt annerledes enn DNA og proteiner fra deres foreldre.
Mye av det tidlige arbeidet i fylogenier, berodde på proteiner. Men å bestemme protein-sekvenser er omstendelig arbeid. Med utvikling av raskere teknikker for å bestemme DNA-sekvenser, ble det mer vanlig å analysere DNA eller RNA som koder for proteiner, heller enn proteinene selv. Omforming av informasjon i DNA sekvenser til proteiner, avhenger av komplekse molekylære strukturer i cellen, kalt ribosomer. De består delvis av ribosomalt RNA (rRNA). Siden 1980 har DNA sekvenser som koder for rRNA forsynt mange av dataene for molekylær fylogeni.
Selv om sammenligning av DNA eller RNA sekvenser er enkelt i teorien, så er det komplekst i praksis. Siden et aktuelt DNA-segment kan inneholde tusener av underenheter, kan det å stille dem opp for sammenligning være tricky. Og ulike oppstillinger kan gi meget ulike resultat. Ikke desto mindre er konklusjoner fra molekylære sammenligninger blitt benyttet i forhold til nøkkel-begivenheter i livets historie. Erfaringene med slike molekylære fylogenier har vært forvirrende, fordi resultatet har vist seg å avhenge ikke bare av hvilken rRNA som analyseres eller endog hvilket laboratorium som utfører sammenligningen. I 1999 skrev evolusjonsbiolog M. Lynch: "Klargjøring av de fylogenetiske relasjoner til dyre-rekkene (phyla) er blitt et vanskelig problem, med analyser basert på ulike gener og endog ulike analyser basert på de samme gener, medfører en diversitet av fylogenetiske trær." (21)
Bilde 1.Molekylær fylogeni -bragte inn noe nytt
Problemene bare fortsatte: I 2002 sammenlignet biolog J.A. Blair og kollegerover 100 protein-sammenstillinger og konkluderte: "Gruppering av rundormer (nematods) med leddyr/insekter (arthropods) er en artefakt som oppsto fra analysen av ett enkelt gen (18s rRNA). Resultatene her foreslår forsiktighet i å foreta dyre-fylogenier basert på analyse av ett eller noen få gener.. Våre resultater indikerer at insekter (arthropods) er genetisk og evolusjonært nærmere mennesker (virveldyr) enn til rundormer (nematods).
i 2004 analyserte forsker ved National Center of Biotechnology Information, Y.I. Wolf og hans kolleger over 500 proteiner i det de benyttet tre ulike fylogenetiske metoder. De konkluderte med at "majoriteten av metodene .. grupperte flua i lag med mennesker til fortrengsel for rundormer." (24) I 2005 analyserte fransk biolog H. Phillippe og kolleger 146 gener og 35 arter, som representerte 12 dyre-rekker. De konkluderte at deres data grupperte leddyr/insekter (arthropods) i lag med rundormer, til fortrengsel for virveldyr. (25) I 2005 benyttet biolog
A. Rokas og kolleger to ulike metoder for å analysere 50 ganer fra 17 dyregrupper. De bemerket at "ulike fylogenetiske kan nå motsigende slutninger med absolutt sikkerhet" og konkluderte at det evolusjonære slektskap mellom dyre-rekkene "forblir uløst." (27)
Selv om fylogenetiske metoder har vist seg problematiske for å redegjøre for relasjoner mellom dyre-rekker, er de mer vellykket i å finne en felles stamfar for alle levende organismer? Nei, faktisk feiler molekylære metoder mer tydelig. I 1980-åra var C.R. Wose pioner på å benyttet genet 18s rRNA for å konstruere et alt-omfattende livets tre og identifisere den universelle felles stamfar. Men ved slutten av 1990-årene hadde seriøse problemer dukket opp. "Da vitenskapsfolk startet å analysere en variasjon av gener fra ulike organismer," skrev molekylær biolog J.A. Lake, R. Jain og M.A. Rivera, fant de at "deres relasjoner til hverandre motsa det evolusjonære livets tre, oppnådd fra rRNA analyser alene." (29) I følge H. Philippe og P. Forterre: " Med flere og flere sekvenser tilgjengelig, viste det seg at de fleste protein-fylogenier motsa hverandre så vel som rRNA-treet." (30)
 I 1998 skrev Wose: "Ingen konsistent organisme-fylogeni har dukket opp fra de mange individuelle protein-fylogenier som er produsert så langt." Han konkluderte at primitive (prokaryote) organismer fikk mange av sine gener og proteiner, ikke ved darwinistisk nedarving med endring, men ved "sideveis genoverføring", der organismer byttet gener med andre organismer. Den universelle stamfaren, skrev han, er ikke en entitet, en ting, men ett fellesskap av komplekse molekyler -en slags ursuppe, fra hvilken ulike slag celler dukket opp uavhengig av hverandre." (31) Omtrent samtidig konkluderte evolusjonsbiolog W.Ford Doolittle ved Dalhouse University, at "sideveis genoverføring blant gamle organismer innebar at molekylære fylogenier ikke kunne bli i stand til å oppdage det sanne livets tre, ikke fordi de benyttet feil metoder eller feil gener, men fordi livets historie ikke kan uttrykkes riktig på treform." Han konkluderte: "Kanskje det ville være lettere og i det lange løp mer produktivt å forlate forsøket på å tvinge" de molekylære data "inn i formen forsynt til Darwin." (32) I stedet for et tre, foreslo Doolittle en vev eller nettverkslignende mønster." (33)
I 1998 skrev Wose: "Ingen konsistent organisme-fylogeni har dukket opp fra de mange individuelle protein-fylogenier som er produsert så langt." Han konkluderte at primitive (prokaryote) organismer fikk mange av sine gener og proteiner, ikke ved darwinistisk nedarving med endring, men ved "sideveis genoverføring", der organismer byttet gener med andre organismer. Den universelle stamfaren, skrev han, er ikke en entitet, en ting, men ett fellesskap av komplekse molekyler -en slags ursuppe, fra hvilken ulike slag celler dukket opp uavhengig av hverandre." (31) Omtrent samtidig konkluderte evolusjonsbiolog W.Ford Doolittle ved Dalhouse University, at "sideveis genoverføring blant gamle organismer innebar at molekylære fylogenier ikke kunne bli i stand til å oppdage det sanne livets tre, ikke fordi de benyttet feil metoder eller feil gener, men fordi livets historie ikke kan uttrykkes riktig på treform." Han konkluderte: "Kanskje det ville være lettere og i det lange løp mer produktivt å forlate forsøket på å tvinge" de molekylære data "inn i formen forsynt til Darwin." (32) I stedet for et tre, foreslo Doolittle en vev eller nettverkslignende mønster." (33)
Bilde 2. Livets tre -slik det sto i 'Origins'
Kontroversen fortsatte: I 2002 foreslo Wose at biologi skulle gå utenom Darwins doktrine om felles avstamning. (35) I 2004 skrev han: "Roten til det universelle treet, er en artefakt som stammet fra å tvinge den evolusjonære kursen inn i en tre-representasjon, når den representasjonen er upassende." (36) I 2004 foreslo W.F. Doolittle og kolleger å erstatte livets tre med en vevs-lignende syntese av livet." (37) og i 2005 anbefalte de at "andre representasjoner enn et tre skulle undersøkes." (38) I mellomtiden fortsatte andre vitenskapsfolk å forsvare hypotesen at det fantes en universell stamfar, men den var kompleks heller enn enkel." (39A)
Det synes klart at molekykær-fylogenien har feilet i å etablere den universelle felles avstamning, og den har feilet i å gi en pålitelig beretning om livets historie. Den har feilet i å spesifisere hvilke evolusjonære relasjoner,.. som eksisterer mellom levende former. Og den har feilet spesielt i å gi en klar indikasjon på hvordan den universelle stamfaren, om den finnes, kan ha sett ut. Var det en organisme, en gruppe av organismer, eller en gruppe av kjemikalier? Det molekylære beviset for felles opphav .. er plaget med usammenhengende og ubesvarte spørsmål. Faktisk må ND antas for å forklare, eller i stigende grad -bortforklare, bevisene.
6. Vestigale strukturer-beste bevis for evolusjon?
I biologi er en vestigal struktur en som utførte en nyttig funksjon i fortiden, men ikke nå lenger. Strukturens fortsatte eksistens synes derfor å være resultat av hva som kunne kalles 'forplantnings-treghet'. Strukturen synes å bli reprodusert fra en generasjon til en annen fordi den er lettere å fortsette å reprodusere enn å kutte den bort og eliminere den for alltid. Det forutsetter at strukturen ikke er skadelig, i hvis tilfelle naturlig seleksjon ville arbeide direkte for å eliminere den. Men biologi synes å presentere oss for vestigale strukturer som verken er skadelige eller nyttig, men bare bæres med for turens skyld.
Hvorvidt en struktur er vestigal i betydning av å ha mistet all funksjonalitet, er ofte en sak til diskusjon. Mange organer som tidligere ble betraktet som vestigale ble senere funnet å ha en funksjon. F.eks. siterer ofte biologi-lærebøker menneskelig 'haleben' som en vestigal struktur. Halebenet består av flere virvler ved slutten av ryggraden. I anatomiske og medisinske lærebøker finner en at halebenet er et viktig kontaktpunkt med muskler som fester seg til undersiden av bekkenet. Lærebøker .. vil beskrive denne og andre funksjoner med menneskelig haleben ganske saklig. Det samme er tilfellet med menneskelig blindtarm. Etter tidligere å ha blitt ansett som vestigal, er blindtarmen nå kjent å være en fungerende komponent i immunsystemet. {Størrelsen på blindtarmen synes å være knyttet til fortæring av cellulose (i blader). Vår lille blindtarm antyder at blader ikke er en viktig del av kostholdet, og er ikke noe høy-tillitsbevis på at vi kommer fra pattedyr som hadde en større blindtarm enn vår. Det samme gjelder det såkalte 'halebenet', som utgjør feste til 9 muskler med tilknytning til nedre del av bekkenet-se Bilde 3. I forbindelse med bekkenben til hvaler og delfiner, er det nå påvist at disse spiller en viktig rolle i forbindelse med reproduksjon (39B) -oversetters tilføyelser: 'The Scientific Approach to Evolution', Rob Stadler, s.64}
Selv om mange vestigale organer ved nærmere undersøkelse er funnet å ha funksjon når alt kommer til alt, så eksisterer det også funksjoner som det ikke kan være tvil om er vestigale, og dermed ubrukelige. Det gjelder f.eks. øyet til visse salamandere eller fisk: 'blinde salamandere' eller 'hulefisk' har bare 'knopper', men ikke virkelige øyne. Disse organismene bebor omgivelser der lyset er totalt fraværende. I et klassisk tilfelle av 'use it or lose it' synes forfedrene til disse organismene på et tidspunkt å ha mistet sine øyne.
Slike vestigale strukturer er helt konsistente med intelligent design, som foreslår at strukturer som opprinnelig ble designet, men så mistet sin funksjon gjennom ulykker eller mangel på bruk. Vestigale strukturer gir også bevis for en begrenset form for devolusjon, som bare forklarer tap av funksjonen og ikke dens opprinnelse. Vestigalitet er i beste fall-evolusjonært sett, en degenerativ form for evolusjon, der pre-eksisterende funksjonelle strukturer endrer seg og taper sin funksjon. Det kan altså ikke gi noe bevis for økende kompleksitet eller tillegg av funksjon- type evolusjon. Det tilbyr ikke innsikt i noen mekanismer som kan være ansvarlig for kreative innovasjoner. Så er spørsmålet hvorvidt det delvis kan bekrefte en felles avstamning?
Bilde 3. Mye folkelig overtro om 'halebenet'
Tilfellet med vitamin-C dannelse i pattedyr gir interessant innsikt i vestigalitet og problemet med å etablere endog begrenset felles avstamning. Selv om de fleste pattedyr kan danne sitt vitamin C, så kan ikke mennesker og marsvin det. (Heller ikke primater generelt eller flaggermus som spiser frukt, kan det). For å danne vitamin C trengs et enzym som kodes av et gen, kjent som GULO. Hva skjedde med det genet i mennesker og marsvin? De har begge ikke-fungerende mutant-kopier av genet. (40) I et tilfelle av 'bruk det eller mist det', har begge arter startet med et fungerende gen, og så mistet det. (Formodentlig fordi dietten deres var rik på C-vitaminer, og det ikke var behov for å produsere det.)
Defekte gener som ikke lenger koder for funksjonelle proteiner, er kjent som pseudogener. Evolusjons-biologer ser disse tilsynelatende vestigale levninger av GULO-genet som bevis for felles avstamning (og som bevis mot Intelligent Design -ID). Men hvorfor skulle disse 'delte feilene' ekistere dersom mennesker ikke stammet fra en felles stamfar delt med andre primater?
Men en studie fra 2003 (Inai et al), som sammenlignet GULO pseudogenet fra marsvin med menneskers, viser mange av de 'delte feilene' mellom de to artene. (41) Mennesker og marsvin er jo ikke såkalte 'søster-arter', blant pattedyrene. Dermed kunne ikke GULO-genet i de to artene stamme fra samme opprinnelige pseudogen, til tross for deres 'delte feil'. Endringene i sekvenser av GULO-genet, må ha inntruffet parallelt. Men om det inntraff uavhengig av hverandre i disse to artene, hvordan kan vi vite at de 'delte feilene' utvist av andre arter (f.eks. sjimpanser og mennesker) kan ha inntruffet parallelt? Inai et al. argumenterer derfor for at GULO-sekvensene kan være utsatte regioner (hot spots), som mer sannsynlig utsettes for erstatninger. (42) Men i det tilfellet kan hva som synes å være 'delte feil', reflektere en økt sannsynlighet for uavhengige erstatninger langs kodende sekvenser, i stedet for felles avstamning. Når vi ser slike såkalt 'delte-feil', så kan det være endringer som oppstår i parallell blant grupper som ikke nødvendigvis stammer fra noen felles stamfar.
I spørsmål om delte feil, er det et poeng først å finne ut hvorvidt det virkelig er feil. Ved liknende koding, er et problem at stille mutasjoner kan endre DNA-sekvensen uten å endre aminosyre-sekvensen i det resulterende proteinet. Det skyldes at flere enn et kodon (triplet av DNA) kan kode for en aminosyre. Det er faktisk mulig for ulike DNA-sekvenser som koder for samme aminosyre sekvens, å resultere i ulike proteiner. David Springer skriver: ribosomet produserer kodoner i ulik hastighet, når kodonet skiller seg bare ved en redundant nukleotide erstatning." Etter som folding av proteinet skjer etter produksjonen, kan det også påvirke måten proteinet folder seg på. Det kan også muligens skyldes at "RNA mollekyler avhengig av spesifikke gen-sekvenser endrer måten proteinet blir prosessert på, etter at ribosomet er ferdig med å produsere det, se her ( referanse 45, bak i boka) Slike mutasjoner, som ikke endrer aminosyresekvensen til resulterende protein, kalles stille (eller synonyme) mutasjoner.
 At proteiner syntetisert med samme aminosyre-sekvens, men ulik DNA-sekvens, kan ha ulike egenskaper er bekreftet ved forskning av Chava Kimchi-Sarfaty ved National Cancer Institute i Bethesda, Maryland. Eksperimenter viste da at "stille mutasjoner" kan danne en merkbar forskjell i assosierte protein produkter, og endre raten der proteiner pumper medisiner fra celler, og derved påvirke resistansen ved visse krefttyper. (44) Slike funn undergraver de 'delte-kodings' argumentene som forutsetter at når gener koder for de samme aminosyrene, produserer de også samme proteinet. Det er ikke tilfellet. Så vestigalitet etablerer ikke entydig felles avstamning. Det har også vært framført argumenter i forhold til retro-transposoner, som vi henviser til her-se slutten. Det er også en mulighet for ennå uoppdagede funksjoner. Dessuten opererer alle disse vestigal-argumentene innenfor samme dyrerekke -phylum). Dermed er det ikke snakk om felles avstamning i vanlig ND ('livets tre') betydning, der hver organisme sporer sin linje tilbake til en felles universal stamfar.
At proteiner syntetisert med samme aminosyre-sekvens, men ulik DNA-sekvens, kan ha ulike egenskaper er bekreftet ved forskning av Chava Kimchi-Sarfaty ved National Cancer Institute i Bethesda, Maryland. Eksperimenter viste da at "stille mutasjoner" kan danne en merkbar forskjell i assosierte protein produkter, og endre raten der proteiner pumper medisiner fra celler, og derved påvirke resistansen ved visse krefttyper. (44) Slike funn undergraver de 'delte-kodings' argumentene som forutsetter at når gener koder for de samme aminosyrene, produserer de også samme proteinet. Det er ikke tilfellet. Så vestigalitet etablerer ikke entydig felles avstamning. Det har også vært framført argumenter i forhold til retro-transposoner, som vi henviser til her-se slutten. Det er også en mulighet for ennå uoppdagede funksjoner. Dessuten opererer alle disse vestigal-argumentene innenfor samme dyrerekke -phylum). Dermed er det ikke snakk om felles avstamning i vanlig ND ('livets tre') betydning, der hver organisme sporer sin linje tilbake til en felles universal stamfar.
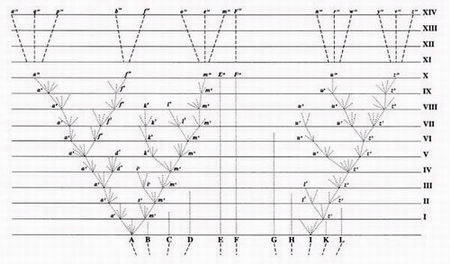
Bilde 4. Haeckels falske tegninger
Selv om argumentene ovenfor skulle innebære en slik avstamning innenfor samme rekke (phylum), så sier det oss ingenting om evolusjonens mekanisme, mye mindre om hvordan nye funksjoner oppsto i første omgang. I forbindelse med vestigale organer er det ikke snakk om å vise evolusjon som en kreativ kraft, i beste fall (for ND) viser de levninger av evolusjon etter at den har gjort sitt. Viktigst er at de er helt i samsvar med evolusjon som en prosess styrt av en konstruerende (designing) intelligens. M.a.o. får ikke vestigalitet design til å forsvinne.
7. Rekapitulasjon
Ang. Rekapitulasjon vil vi henvise til tidligere lenker herfra. Etter 100 år med feilinformasjon i lærebøker om Haeckels fostre, så er det på tide med en smekk til dem som holder på med dette: Stadiet som Haeckel kalte det første er faktisk midtveis i embryo-utviklingen. Faktisk frembyr de første stadiene slående ulikheter, forut for de stadiene der Haeckel overdrev likhetene. Darwins "sterkeste enkelt-gruppe fakta", består dermed i et klassisk tilfelle av at man fordreier vitenskapelige bevis, for å få det til å passe med en teori. Teorien om rekapitulasjon tvinger embryo-bevis inn i ND-form. Ved å gjøre det ødelegger rekapitulasjonsteorien vår forståelse av både embryologi og evolusjon.
 8. Felles design -avstamning eller begge?
8. Felles design -avstamning eller begge?
Likheter (strukturelle og funksjonelle) kan forklares både ved felles design og/eller felles avstamning, og i prinsippet ved begge. I fravær av design, som i filosofisk naturalisme, så blir felles avstamning eneste alternativ. Men om design er tilbake på banen, kan ikke felles avstamning tas for gitt. Dermed er ikke påvisning av likheter tilstrekkelig til å forklare årsaken til disse. Felles design kan alene være ansvarlig for de lignende (strukturelle og funksjonelle) trekkene som går igjen i biologien. I analogi-forklaringer kan en illustrere med felles-subrutiner, som benyttes i ulike dyrs embryo-program. Vi har henvist til tolkningsmuligheter i forbindelse med nettverks-mønster av likheter. Vi henviser her bare til hva som ble sagt i forbindelse med pungdyr -lenke. Flyge-evne skal i følge ND ha inntruffet uavhengig av hverandre 4 ganger: for fugler, insekter, pattedyr (flaggermus) og pterosaurus (utdødde flygende reptiler).
Både spor av design og spor av ulykker og feil er til stede i biologien. Ulykker gjennom historien kan re-arrangere, modifisere og også bryte funksjonsevne i design-moduler. Når de en gang er introdusert, kan de føres videre: Likheter som skyldes tap av funksjonsevne i gjennom livets historie, 'forplantnings-treghet', trekk som blir med på lasset, fordi seleksjonsprosessen ikke siler ut det som reproduseres.
Bilde 5. Reell motsetning her
Prinsipielt er det viktig ikke å havne i falske dilemmaer, som er valg mellom to alternativer som ikke er fullstendig akseptable, men som til sammen synes å være gjensidig utelukkende og fullstendige. I prinsippet er et slikt falskt dilemma: felles design kontra felles avstamning. Vi kan tenke på livet som hierarkisk arrangerte design-moduler, som i løpet av livets historie har opprettholdt betydelige evolusjonære endringer, gjennom naturlige prosesser så vel som styring av en veiledende intelligens. Akkurat hvor mye, når og hvordan forblir inntil videre et åpent spørsmål. Vi kan selvsagt i prinsippet også ha felles design uten felles avstamning og v.v.
Det bringer oss til et sant dilemma: Valget mellom ID og filosofisk naturalistisme. Sistnevnte omfavner ikke bare felles avstamning, men avviser også ethvert virkelig design. ID til sammenligning hevder at biologisk design er virkelig og empirisk oppdagbart, uansett hvorvidt det opptrer innen en evolusjonær prosess eller i diskrete uavhengige steg. Tilhengere av ID kan holde på ulike grader av avstamning, men vil alltid holde på en åpning for design. Sammen kan disse to forklare lignende (strukturelle og funksjonelle) trekk i biologien. Det presserende spørsmål er hvorvidt felles avstamning atskilt fra felles design kan gjøre det. Biologiske resultater indikerer i økende grad at det ikke kan det.
Referanser:
21. M. Lynch, The Age and Relationship of the Major Animal Phyla, Evolution 43 (1999): 319-325
24. Yuri I. Wolf, et al, Coelomata and Not Ecdysozoa: Evidence fra Genome-Wide Phylogenetic Analysis, Genome Research 14 (2004): 2936. http://genome.cshlp.org/content/14/1/29.full.pdf
25. H. Phillippe et al. Multigene Analysis of Bilaterian Animals Cooroborate the Monophyly of Ecdysozoa, Lophotrochozoa and Protostomia, Molecular Biology and Evolution, 22 (2005): 1246-1253
27. A. Rokas et al, Animal Evolution and the Molecular Signature of Radiations compressed in Time, Science 310(2005):1933-1938
29. Darwin, Origin of Species, 484
30. H. Phillippe et al, The Rooting of the Universal Tree of Life Is not Reliable, Journal of Molecular Evolution 49 (1999):509-523
31. C. Woese, The Universal Ancestor, Proceedings of the National Academy of Sciences USA 95 (1998):6854-6859
32. W. F. Doolittle, Phylogenetic Classification and the Universal Tree, Science 284 (1999): 2124-2128
33. W. F. Doolittle, Uprooting the Tree of Life, scientific American 228 (Febr. 2000): 90-95
35. C. Woese, On the Evolution of Cells, Proceedings of the National Academy of Science USA 99 (2002): 8742-8747
36. C.R. Woese, A new Biology for a New Century, Microbiology and Molecular Biology reviews 68 (2004): 173-186
37. W.F. Doolittle, If the Tree of Life Fell, Would We REconize the Sound? 119-133 i Microbiological Phylogeny and Evolution: Concepts and Controversies (New York: Oxford University Press, 2005), 131
38. E. Bapteste et al, Do Orthologous Gene Phylogenies Really Support Tree-Thinking? BioMed Central Evolutionayr Biology 5 (2005): 33 http://www.biomedcentral.com/content/pdf/1471-2148-5-33.pdf
39.A. M.C.Rivera and J.A.Lake, The RIng of Life Provides Evidence for a Genome Fusion Origin of Eukaryotes, Nature 431 (2004). 152-155
39.B. Dines, J.P, Sexual selection targets cetacean pelvic bones. Evolution 2014; 68: 3296-3306
40. M. Nishikimi et al, Guinea pigs Posess a Highly Mutated Gene for L-Gulono-Gamma-Lactone Oxidase, Journal of Biological Chemistry 269 (1994): 13685-13688
41. Y. Inai et al, The Whole Structure of the Human Non-funkctional L-Gulono-gamma-lactone Oxidase Gene - Journal of Nutrition Science and Vitaminology (Tokyo) 49 (2003): 315-319
42. Inai et al skriver: "Assuming an equal chance of substitution ... was calculated to be 1.84*10 opphøyd i -12. This extremely small probability indicates the precense of many mutational hot spots in the sequences" (317)
Oversettelse og bilder ved Asbjørn E. Lund